






Metabolismo do lago

Content
Conceito
As estimativas do metabolismo do lago normalmente dependem da medição de oxigênio dissolvido ou dióxido de carbono, ou medições de um traçador de carbono ou oxigênio para estimar a produção e o consumo de carbono orgânico. O oxigênio é produzido e o dióxido de carbono consumido através da fotossíntese e o oxigênio é consumido e o dióxido de carbono produzido através da respiração. Aqui, a matéria orgânica é simbolizada pela glicose, embora as espécies químicas produzidas e respiradas através dessas reações variem amplamente.

Fotossíntese: 6 c o 2 + 6 h 2 o → l i g h t c 6 h 12 o 6 + 6 o 2 {\ displayStyle 6co_ {2} + 6h_ {2} o {\ xrightArrow [{}] {luz}} c_ {6} H_ {12} O_ {6}+6o_ {2}}
Respiração: C 6 H 12 O 6 + 6 O 2 → 6 C O 2 + 6 H 2 O {\ DisplayStyle C_ {6} H_ {12} O_ {6} + 6o_ {2} {\ XrighTarrow [{}] {} } 6co_ {2}+6h_ {2} o}
A fotossíntese e a produção de oxigênio ocorre apenas na presença de luz, enquanto o consumo de oxigênio via respiração ocorre tanto na presença quanto na ausência de luz. Os termos do metabolismo do lago incluem:
GPP - gross primary production (e.g. total photosynthesis)R - total respiration ( R h + R a ) {\displaystyle (R_{h}+R_{a})} R h {\displaystyle R_{h}}
R h {\displaystyle R_{h}}  - heterotrophic respiration R a {\displaystyle R_{a}}
- heterotrophic respiration R a {\displaystyle R_{a}}  - autotrophic respirationNEP - net ecosystem production = GPP - RNPP - net primary production = GPP - R a {\displaystyle R_{a}}
- autotrophic respirationNEP - net ecosystem production = GPP - RNPP - net primary production = GPP - R a {\displaystyle R_{a}} Técnicas de medição
A estimativa do metabolismo do lago requer processos de aproximação que influenciem a produção e o consumo de carbono orgânico por organismos dentro do lago. Mudanças cíclicas em escala diária ocorrem na maioria dos lagos na Terra porque a luz solar está disponível para fotossíntese e produção de novo carbono apenas para uma parte do dia. Os pesquisadores podem aproveitar esse padrão de diel para medir as taxas de mudança no próprio carbono ou mudanças nos gases dissolvidos, como dióxido de carbono ou oxigênio que ocorrem em escala diária. Embora as estimativas diárias do metabolismo sejam mais comuns, o metabolismo inteiro pode ser integrado por períodos de tempo mais longos, como taxas sazonais ou anuais, estimando um orçamento de carbono de lago inteiro. As seções a seguir destacam as maneiras mais comuns de estimar o metabolismo do lago em uma variedade de escalas temporais e espaciais e repassar algumas das suposições de cada um desses métodos.

Métodos de água livre
A medição de alterações de diel nos gases dissolvidos dentro do lago, também conhecida como método de "água livre", tornou-se rapidamente o método mais comum de estimar o metabolismo do lago, uma vez que a ampla adoção de sensores autônomos usados para medir oxigênio dissolvido e dióxido de carbono em água . O método de água livre é particularmente popular, pois muitas estimativas diárias do metabolismo do lago podem ser coletadas de forma relativamente barata e podem fornecer informações sobre regimes metabólicos durante períodos de tempo difícil de observar, como durante os eventos de tempestade. Alterações medidas no oxigênio dissolvido e dióxido de carbono dentro de um lago representa a soma de todo o metabolismo organismo de bactérias a peixes, após a contabilização de alterações abióticas nos gases dissolvidos. As alterações abióticas nos gases dissolvidos incluem trocas de gases dissolvidos entre a atmosfera e a superfície do lago, arrastamento vertical ou horizontal da água com concentrações diferentes (por exemplo, água de baixo oxigênio abaixo de um termoclina de um lago), ou importação e exportação de gases dissolvidos de riachos de entrada ou a uma saída do lago. Alterações abióticas nos gases dissolvidas podem dominar mudanças de gases dissolvidos se o lago tiver uma baixa taxa metabólica (por exemplo, lago oligotrófico, dia nublado), ou se houver um grande evento que causa fatores abióticos excedem bióticos (por exemplo, eventos de vento causando mistura e entrada de água com baixa oxigenada). Os sinais bióticos em gases dissolvidos são mais evidentes quando o sol está brilhando e a fotossíntese está ocorrendo, resultando na produção de oxigênio dissolvido e consumo de dióxido de carbono. A conversão da energia solar em energia química é denominada produção primária bruta (GPP) e a dissipação dessa energia através da oxidação biológica de carbono é denominada respiração do ecossistema (ER). As medições de alta frequência (por exemplo, 10 minutos) de oxigênio dissolvido ou dióxido de carbono podem ser traduzidas em estimativas de GPP, ER e a diferença entre os dois denominados ecossistemas líquidos (NEP), ajustando os dados de alta frequência a modelos de modelos de Metabolismo do lago. A equação que governa para estimar o metabolismo do lago a partir de um único sensor localizado na camada mista superior medindo o oxigênio dissolvido é:
Do/t = gpp-er+f
Onde f é o fluxo de gases entre o lago e a atmosfera. Termos adicionais de fluxo de gás abiótico podem ser adicionados se esses fluxos abióticos forem considerados significativos para um lago (por exemplo, mistura de eventos, entrada de gases de fluxo). A troca gasosa atmosférica (F) raramente é diretamente medida e normalmente modelada pela estimativa da turbulência da superfície do lago a partir da mistura orientada pelo vento e convectivo. Na maioria das vezes, F é estimado a partir de medições de velocidade do vento e pressão atmosférica, e modelos diferentes para estimar F podem resultar em estimativas significativamente diferentes das taxas metabólicas do lago, dependendo do lago de estudo. Presume -se que a produção primária bruta seja zero durante a noite devido a baixa ou nenhuma luz e, portanto, o ER pode ser estimado a partir de alterações noturnas no oxigênio dissolvido (ou dióxido de carbono) após contabilizar alterações abióticas no oxigênio dissolvido. A produção primária bruta pode ser estimada, assumindo que o ER é igual durante o dia e a noite e responde a alterações dissolvidas de oxigênio durante o dia, no entanto, essa suposição pode não ser válida em todos os lago.
Extrair uma alta relação sinal / ruído é essencial para obter boas estimativas do metabolismo do lago da técnica de água livre, e há opções que um pesquisador precisa fazer antes dos dados da coleta e durante as análises de dados para garantir estimativas precisas. Localização da coleta de gás dissolvida (normalmente na camada mista de superfície), número de sensores vertical e horizontalmente, frequência e duração da coleta de dados e métodos de modelagem precisam ser considerados.
Free-water metabolism modeling techniquesAs técnicas de medição de águas livres exigem modelos matemáticos para estimar as métricas do metabolismo do lago a partir de medições de gás dissolvidas de alta frequência. Esses modelos variam em complexidade, desde modelos algébricos simples até modelagem integrada a profundidade usando técnicas estatísticas mais avançadas. Várias técnicas estatísticas foram usadas para estimar GPP, R e NEP ou parâmetros relacionados a esses termos do metabolismo.
Métodos de garrafa clara e escura
O método de garrafa claro e escuro usa o mesmo conceito que o método de água livre para estimar as taxas de metabolismo - o GPP ocorre apenas durante o dia com energia solar enquanto o ER ocorre na presença e na ausência de luz. Esse método incuba a água do lago em duas garrafas separadas, uma que é clara e exposta ao regime de luz natural ou artificial e outro que é fechado da luz envolvendo a garrafa em papel alumínio, tinta ou outro método. Alterações na fixação de carbono ou gases dissolvidas são então medidas ao longo de um determinado período de tempo (por exemplo, várias horas a um dia) para estimar a taxa de metabolismo para profundidades específicas do lago ou uma coluna de água do lago integrada. A fixação do carbono é medida pela injeção de isótopo de carbono radioativo 14C em garrafas claras e escuras e amostrando as garrafas ao longo do tempo - as amostras são filtradas no papel de filtro e a quantidade de 14C incorporada nas células de algas (e bacterianas) é estimada medindo amostras em A contador de cintilação. A diferença entre a garrafa clara e escura 14C pode ser considerada a taxa de produtividade primária; No entanto, devido à captação não fotoossintética do CO2, há debate sobre se as garrafas escuras devem ser usadas com o método 14C ou se apenas uma garrafa de luz e uma garrafa tratada com o algicida DCMU devem ser usadas. As taxas de mudança de gases dissolvidos, dióxido de carbono ou oxigênio, precisam de garrafas claras e escuras para estimar as taxas de produtividade e respiração.
Métodos de orçamento de carbono de lago inteiro
Provavelmente, o método mais trabalhoso de estimativa de uma métrica do metabolismo do lago é medindo todos os insumos e saídas de carbono orgânico ou inorgânico em um lago durante uma temporada ou ano, também conhecido como orçamento de carbono inteiro. A medição de todas as entradas e saídas de carbono de e para um lago pode ser usada para estimar a produção líquida do ecossistema (NEP). Como o NEP é a diferença entre produção primária bruta e respiração (NEP = GPP - R), ele pode ser visto como a conversão biológica líquida de carbono inorgânico em carbono orgânico (e vice -versa) e, portanto, pode ser determinado através da massa de lago inteiro Equilíbrio de carbono inorgânico ou orgânico. O NEP avaliado por meio de carbono inorgânico (IC) ou orgânico (OC) pode ser estimado como:
N e p o c = e o c + s o c-i o c {\ displaystyle nep_ {oc} = e_ {oc} + s_ {oc} -i_ {oc}}
N e p i c = i c-s i c-e i c {\ displaystyle nep_ {ic} = i_ {ic} -s_ {ic} -e_ {ic}}
onde E é a exportação de CO através do transporte fluvial e IC através do transporte fluvial e gás de carbono (por exemplo, CO2, CH4) entre a superfície do lago até a atmosfera; S é armazenamento na coluna de sedimentos e água do lago para coluna de OC e água para IC; e eu é a entrada de OC e IC das vias fluviais, vizinhas de zonas úmidas e transportadas pelo ar (por exemplo, deposição atmosférica, queda de ninhada). Um lago que recebe mais OC da bacia hidrográfica do que exporta a jusante ou se acumula na coluna de água e nos sedimentos (IOC> EOC + SOC) indica que houve conversão líquida de OC em IC dentro do lago e, portanto, é heterotrófica líquida (NEP negativa) . Da mesma forma, um lago que acumula e exporta mais IC do que foi recebido da bacia hidrográfica (sic + eic> iic) também indica a conversão líquida de OC em IC dentro do lago e, portanto, é heterotrófica líquida.
Métodos de metabolismo bentônico
Embora o método de água livre provavelmente contenha algum sinal metabólico bentônico, o isolamento da contribuição bentônica para o metabolismo do lago inteiro requer métodos específicos bentônicos. Análoga aos métodos de garrafa claro e escura descritos acima, os núcleos de sedimentos do lago podem ser coletados e alterações no oxigênio dissolvido ou na fixação de carbono podem ser usadas para estimar taxas de produtividade e respiração primária. Métodos relativamente novos descrevem o isolamento da interface de água de sedimentos com cúpulas transparentes e medem alterações no oxigênio dissolvido in situ, que é um híbrido entre o método de água livre e o método da garrafa de escuro leve. Esses métodos de câmara bentônica in situ permitem estimativa relativamente fácil de vários dias do metabolismo bentônico, o que ajuda o pesquisador a determinar como o metabolismo bentônico muda com padrões climáticos variados e características do lago.
Suposições
O local extrapolador ou medições específicas de profundidade para todo o lago pode ser problemático, pois pode haver uma variabilidade metabólica significativa, vertical e horizontalmente dentro de um lago (consulte a seção de variabilidade). Por exemplo, muitos estudos do metabolismo do lago têm apenas uma única estimativa epilimnética do metabolismo; no entanto, isso pode superestimar as características metabólicas sobre o lago, como a NEP, dependendo da profundidade da profundidade da camada mista para a profundidade da extinção da luz. As estimativas diárias de metabolismo diário por períodos de tempo mais longos podem ajudar a superar alguns desses problemas de extrapolação do local, mas é preciso considerar cuidadosamente as implicações das estimativas metabólicas e não sobre as medições extrapoladas.
Relação com constituintes
A taxa metabólica organisma, ou a taxa na qual os organismos assimilam, transformam e gastam energia, são influenciados por alguns constituintes -chave, a saber, leve, nutrientes, temperatura e matéria orgânica. A influência desses constituintes no metabolismo organismo governa o metabolismo em toda a escala do lago e pode ditar se um lago é uma fonte líquida ou pia de carbono. Na seção a seguir, descrevemos a relação entre esses constituintes-chave e o metabolismo organismo e no nível do ecossistema. Embora as relações entre organismos e constituintes descritos aqui estejam bem estabelecidos, os efeitos interagidos dos constituintes nas taxas metabólicas, desde organismos até ecossistemas do lago, dificultam a previsão de mudanças no metabolismo nos lagos ou dentro dos lagos ao longo do tempo. Muitos desses efeitos complexos de interação serão discutidos na seção de variabilidade espacial e temporal.
Temperatura

A temperatura é um forte fator de controle nas taxas de reação bioquímica e atividade biológica. A temperatura ideal varia entre os organismos aquáticos, pois alguns organismos são mais adaptados ao frio, enquanto outros preferem habitats mais quentes. Existem casos raros de extrema tolerância térmica em lagos antárticos hipersalinos (por exemplo, Don Juan Pond) ou fontes termais (por exemplo, gêiserer de mosca); No entanto, a maioria dos organismos do lago na Terra reside em temperaturas variando de 0 a 40 graus C. As taxas metabólicas normalmente escalam exponencialmente com a temperatura, no entanto, a energia de ativação para produtividade e respiração primária diferem frequentemente, com a fotossíntese com menor energia de ativação do que a respiração aeróbica. Essas diferenças nas energias de ativação podem ter implicações para o equilíbrio metabólico líquido nos ecossistemas do lago à medida que o clima se aquece. Por exemplo, Scharfenberger et al. (2019) mostram que o aumento da temperatura da água resultante das mudanças climáticas pode mudar os lagos de serem líquidos autotróficos para heterotróficos devido a diferenças na energia de ativação, no entanto, a temperatura na qual eles alternam depende da quantidade de nutrientes disponíveis.
Nutrientes
A quantidade de material disponível para assimilação em células organismos controla a taxa de metabolismo no nível do ecossistema celular ao lago. Nos lagos, o fósforo e o nitrogênio são os nutrientes limitantes mais comuns da produção primária e respiração do ecossistema. O trabalho fundamental sobre a relação positiva entre a concentração de fósforo e a eutrofização do lago resultou em legislação que limitava a quantidade de fósforo em detergentes da lavanderia, entre outros regulamentos. Embora o fósforo seja frequentemente usado como preditor da produtividade do ecossistema do lago e do excesso de fósforo como um indicador de eutrofização, muitos estudos mostram que o metabolismo é co-limitado por fósforo e nitrogênio ou nitrogênio sozinho. O equilíbrio entre fósforo, nitrogênio e outros nutrientes, denominado estequiometria ecológica, pode ditar as taxas de crescimento organismo e metabolismo do lago inteiro através dos requisitos celulares desses nutrientes essenciais mediados por características da história da vida. Por exemplo, os cladoceranos de crescimento rápido têm uma razão de nitrogênio para fósforo muito menor (N: P) do que os copépodes, principalmente devido à alta quantidade de RNA rico em fósforo em suas células usadas para um rápido crescimento. Os cladoceranos que residem em lagos com altas taxas N: P em relação à estequiometria corporal cladocerana serão limitadas em crescimento e metabolismo, tendo efeitos no metabolismo do lago inteiro. Além disso, os efeitos em cascata das manipulações da web alimentar podem causar mudanças na produtividade das mudanças na estequiometria de nutrientes. Por exemplo, a adição do piscívoro pode reduzir a pressão da predação sobre os cladocerantes n: p baixo de crescimento rápido, que aumentam rapidamente a população, retêm o fósforo em suas células e podem fazer com que um lago se torne fósforo limitado, reduzindo consequentemente a produtividade primária do lago inteiro.
Leve
A energia solar é necessária para converter dióxido de carbono e água em matéria orgânica, também conhecida como fotossíntese. Assim como a temperatura e os nutrientes, diferentes algas têm diferentes taxas de resposta metabólica ao aumento da luz e também diferentes condições de luz ideais para o crescimento, pois algumas algas são mais adaptadas para ambientes mais escuros, enquanto outros podem superar em condições mais leves. A luz também pode interagir com os nutrientes para afetar a resposta de produtividade de algas específicas da espécie ao aumento da luz. Essas diferentes respostas no nível organismo se propagam para influenciar o metabolismo no nível do ecossistema. Mesmo em lagos de baixo nutriente, onde se espera que os nutrientes sejam o recurso limitador para a produtividade primária, a luz ainda pode ser o recurso limitador, que tem efeitos negativos em cascata em níveis tróficos mais altos, como a produtividade dos peixes. A variabilidade na luz em diferentes zonas do lago e dentro de um lago através do tempo cria manchas na produtividade espacial e temporalmente.
Além de controlar a produtividade primária, a luz solar também pode influenciar as taxas de respiração por matéria orgânica parcialmente oxidante, o que pode facilitar a quebra de bactérias e converter em dióxido de carbono. Essa fotooxidação parcial aumenta essencialmente a quantidade de matéria orgânica disponível para mineralização. Em alguns lagos, a fotooxidação completa ou a fotooxidação parcial pode ser responsável pela maioria da conversão da matéria orgânica para a inorgânica, no entanto, a proporção à respiração bacteriana varia muito entre os lagos.
Carbono organico
Os consumidores primários e secundários em lagos requerem matéria orgânica (de plantas ou animais) para manter a função organisma. A matéria orgânica, incluindo folhas de árvores, matéria orgânica dissolvida e algas, fornece recursos essenciais a esses consumidores e, no processo, aumentam as taxas de respiração do ecossistema do lago na conversão da matéria orgânica em crescimento celular e manutenção organisma. Algumas fontes de matéria orgânica podem afetar a disponibilidade de outros constituintes. Por exemplo, a matéria orgânica dissolvida geralmente escurece a água do lago, o que reduz a quantidade de luz disponível no lago, reduzindo assim a produção primária. No entanto, aumentos na carga de matéria orgânica em um lago também podem aumentar os nutrientes associados à matéria orgânica, o que pode estimular a produção e a respiração primária. O aumento do carregamento de matéria orgânica dissolvida pode criar compensações entre o aumento da limitação da luz e a liberação da limitação de nutrientes. Essa troca pode criar relações não lineares entre a produção primária do lago e o carregamento de matéria orgânica dissolvida com base em quanto nutrientes estão associados à matéria orgânica e com que rapidez a matéria orgânica dissolvida bloqueia a luz na coluna de água. Isso ocorre porque em baixas concentrações de matéria orgânica dissolvida à medida que a concentração de matéria orgânica dissolvida aumenta, o aumento dos nutrientes associados aumenta a GPP. Mas, à medida que a matéria orgânica dissolvida continua aumentando, a redução na luz do escurecimento da água do lago suprime o GPP à medida que a luz se torna o recurso limitador para a produtividade primária. As diferenças na magnitude e na localização do GPP máximo em resposta ao aumento da carga do DOC são levantadas a surgir com base na proporção de documentos para nutrientes que entram no lago, bem como no efeito do clima de LAKE LAKE. O escurecimento da água do lago também pode trocar regimes térmicos dentro do lago, pois as águas mais escuras geralmente significam que as águas mais quentes permanecem no topo do lago, enquanto as águas mais frias estão no fundo. Essa mudança na distribuição de energia térmica pode afetar as taxas de produtividade pelágica e bentônica (veja a temperatura acima) e alterar a estabilidade da coluna de água, com impactos na distribuição vertical dos nutrientes, tendo efeitos na distribuição vertical das taxas metabólicas.

Outros constituintes
Outros constituintes do lago podem influenciar as taxas metabólicas do lago, incluindo concentração de CO2, pH, salinidade e sílica, entre outros. O CO2 pode ser um recurso limitador (ou co-limitar junto com outros nutrientes) para a produtividade primária e pode promover flores de fitoplâncton mais intensas. Algumas espécies de algas, como os crisófitos, podem não ter mecanismos de concentração de carbono ou a capacidade de usar bicarbonato como fonte de carbono inorgânico para a fotossíntese; portanto, os níveis elevados de CO2 podem aumentar suas taxas de fotossíntese. Durante as flores de algas, o CO2 dissolvido elevado garante que o CO2 não seja um recurso limitante para o crescimento, uma vez que aumentos rápidos na produção esgotam o CO2 e aumentam o pH. Alterações no pH em escalas de tempo curto (por exemplo, sub-diário) de picos na produtividade primária podem causar reduções de curto prazo no crescimento e respiração bacteriana, mas em escalas de tempo mais longas, as comunidades bacterianas podem se adaptar ao pH elevado.
A salinidade também pode causar alterações nas taxas metabólicas dos lagos por meio de impactos de salinidade nas taxas metabólicas individuais e na composição da comunidade. As taxas metabólicas do lago podem ser correlacionadas positivas ou negativamente com a salinidade devido a interações de salinidade com outros fatores de motorista do metabolismo do ecossistema, como taxas de descarga ou secas. Por exemplo, Moreira-Turcq (2000) descobriu que o excesso de precipitação sobre a evaporação causou salinidade reduzida em uma lagoa costeira, aumento da carga de nutrientes e aumento da produtividade primária pelagica. A relação positiva entre produtividade primária e salinidade pode ser um indicador de alterações na disponibilidade de nutrientes devido ao aumento das entradas. No entanto, o aumento da salinidade dos sais rodoviários pode causar toxicidade em alguns organismos dos lagos, e os casos extremos de aumentos de salinidade podem restringir a mistura do lago, o que pode mudar a distribuição das taxas de metabolismo em toda a coluna de água do lago.
Variabilidade espacial e temporal
As taxas metabólicas em lagos e reservatórios são controladas por muitos fatores ambientais, como disponibilidade de luz e nutrientes, temperatura e regimes de mistura de colunas de água. Assim, as alterações espaciais e temporais nesses fatores causam variabilidade espacial e temporal nas taxas metabólicas, e cada um desses fatores afeta o metabolismo em diferentes escalas espaciais e temporais.
Variação espacial nos lagos
Contribuições variáveis de diferentes zonas do lago (isto é, litoral, limnética, bentônica) para o metabolismo inteiro do lago depende principalmente da maquiagem na biomassa de algas e bacterianas e disponibilidade de luz e nutrientes. Em termos de organismos que contribuem para o metabolismo em cada uma dessas zonas, o metabolismo limnético é dominado pelo fitoplâncton, zooplâncton e metabolismo bacteriano, com baixa contribuição de epífitos e peixes. O metabolismo bentônico pode receber grandes contribuições de macrófitas, macro e microalgas, invertebrados e bactérias. O metabolismo bentônico é geralmente mais alto em zonas litorivas rasas, ou em lagos rasos de águas claras, nas quais a luz chega ao fundo do lago para estimular a produção primária. Nos lagos escuros ou turbidos, a produção primária pode ser restrita a águas mais rasas e a respiração aeróbica pode ser reduzida ou inexistente em águas mais profundas devido à formação de zonas profundas anóxicas.
O grau de heterogeneidade espacial nas taxas metabólicas dentro de um lago depende da morfometria do lago, características de bacia hidrográfica (por exemplo, diferenças no uso da terra durante toda a bacia hidrográfica e insumos dos riachos) e processos hidrodinâmicos. Por exemplo, lagos com processos hidrodinâmicos mais intensos, como mistura vertical e lateral fortes, são mais lateralmente e verticalmente homogêneos em relação às taxas metabólicas do que os lagos altamente estratificados. Por outro lado, lagos com áreas mais desenvolvidas litoral têm maior heterogeneidade metabólica lateralmente do que os lagos com uma forma mais circular e baixas proporções de áreas litororais rasas.
A atenuação da luz ocorrendo em toda a coluna de água, em combinação com estratificação térmica e química e turbulência acionada por vento ou convectivo, contribui para a distribuição vertical de nutrientes e organismos na coluna de água. Nos lagos estratificados, a matéria orgânica e os nutrientes tendem a estar mais concentrados em camadas mais profundas, enquanto a luz está mais disponível em camadas mais rasas. A distribuição vertical da produção primária responde a um equilíbrio entre a disponibilidade de luz e nutrientes, enquanto a respiração ocorre mais independentemente da luz e dos nutrientes e mais homogeneamente com profundidade. Isso geralmente resulta em um forte acoplamento da produção primária bruta (GPP) e respiração do ecossistema (ER) nas camadas da superfície do lago, mas no acoplamento mais fraco em profundidades maiores. Isso significa que as taxas de ER dependem fortemente da produção primária em camadas mais rasas, enquanto em camadas mais profundas se torna mais dependente de uma mistura de matéria orgânica de fontes terrestres e sedimentação de partículas de algas e matéria orgânica produzida em camadas rasas. Nos lagos com baixa concentração de nutrientes nas águas da superfície e com penetração de luz abaixo da camada mista, a produção primária é maior nas profundidades intermediárias, onde há luz suficiente para a fotossíntese e maior disponibilidade de nutrientes. Por outro lado, os lagos polimíticos baixos transparentes têm maior produção primária em camadas próximas à superfície, geralmente com um equilíbrio autotrófico líquido (GPP> ER) entre produção e respiração primária.
Lateralmente, a heterogeneidade nos lagos é impulsionada por diferenças nas taxas metabólicas nas zonas limnéticas de água aberta e nas zonas litornas mais dominadas por bentônicas. As áreas litorais são geralmente mais complexas e heterogêneas, em parte devido à sua proximidade com o sistema terrestre, mas também devido ao baixo volume de água e alta razão de volume de sedimentos / água. Assim, as zonas do litoral são mais suscetíveis a mudanças de temperatura, insumos de nutrientes e matéria orgânica da paisagem e entradas de rios, mistura de cisalhamento do vento e ação das ondas, sombreamento da vegetação terrestre e ressuspensão dos sedimentos (Figura 1). Além disso, as zonas do litoral geralmente têm maior complexidade de habitat devido à presença de macrófitas, que servem como abrigo, viveiro e local de alimentação para muitos organismos. Consequentemente, as taxas metabólicas nas áreas do litoral geralmente têm alta variabilidade a curto prazo e são tipicamente maiores que as taxas metabólicas limnéticas.
Variação espacial entre lagos
Além da variabilidade espacial nos lagos, as taxas metabólicas do lago inteiro e seus motoristas também diferem entre os lagos. Cada lago tem um conjunto único de características, dependendo de sua morfometria, propriedades de captação e características hidrológicas. Essas características afetam as condições do lago, como cor da água, temperatura, nutrientes, matéria orgânica, atenuação da luz, mistura vertical e horizontal, com efeitos diretos e indiretos no metabolismo do lago.
À medida que os lagos diferem no status de seus constituintes (por exemplo, luz, nutrientes, temperatura e matéria orgânica), existem diferenças emergentes na magnitude e variabilidade das taxas metabólicas entre os lagos. Na seção anterior (relação com os constituintes), discutimos os padrões esperados de taxas metabólicas em resposta à variabilidade nesses constituintes influentes. Aqui, discutiremos como o metabolismo inteiro varia entre os lagos devido a diferenças nesses constituintes, mediados por diferenças na morfometria do lago, propriedades de captação e tempo de permanência na água.
A morfometria do lago (por exemplo, tamanho e forma do lago) e propriedades de captação (por exemplo, uso da terra, área de drenagem, clima e características geológicas) determinam o fluxo de entradas externas de matéria orgânica e nutrientes por unidade de volume de água do lago. À medida que a proporção entre o tamanho da bacia e o volume da água do lago (taxa de drenagem) aumenta, o fluxo de nutrientes e a matéria orgânica da paisagem terrestre circundante geralmente aumenta. Ou seja, pequenos lagos com bacias hidrográficas relativamente grandes receberão mais insumos externos de nutrientes e matéria orgânica por unidade de volume do lago do que grandes lagos com bacias hidrográficas relativamente pequenas, aumentando assim as taxas primárias de produção e respiração. Em lagos com pequena taxa de drenagem (isto é, uma grande área de superfície relativa do lago em relação à área de captação), os processos metabólicos devem ser menos dependentes de entradas externas provenientes da bacia hidrográfica circundante. Além disso, pequenos lagos são menos expostos à mistura acionada pelo vento e geralmente têm mais informações sobre matéria orgânica terrestre, que geralmente resultam em profundidades de mistura mais rasas e na atenuação da luz aprimorada, limitando assim a produção primária a porções superiores de pequenos lagos. Considerando lagos com propriedades de captação semelhantes, pequenos lagos são geralmente mais heterotróficos líquidos (GPP

Propriedades de captação, nomeadamente cobertura da terra, uso da terra e características geológicas, influenciam o metabolismo do lago através de seu impacto na qualidade da matéria orgânica e dos nutrientes que entram no lago, bem como a exposição ao vento. A qualidade da matéria orgânica pode afetar a atenuação da luz e, juntamente com a exposição ao vento, pode influenciar a distribuição de calor e luz em toda a coluna de Water Lake. Os lagos em paisagens dominados pela agricultura apresentam maiores insumos de nutrientes e insumos de matéria orgânica mais baixos em comparação com lagos com taxa de drenagem semelhante, mas em paisagens dominadas por florestas. Assim, espera-se que os lagos em paisagens dominadas pela agricultura tenham taxas de produção primárias mais altas, mais flores de algas e biomassa de macrófitos excessivos em comparação com lagos em paisagens dominadas pela floresta (Figura). No entanto, os efeitos do tamanho da captação e do tipo de captação são complexos e interativos. Os lagos florestais relativamente pequenos são mais sombreados e protegidos da exposição ao vento e também recebem grandes quantidades de matéria orgânica alóctone. Assim, pequenos lagos arborizados são geralmente mais húmicos com uma camada mista rasa e penetração de luz reduzida. Os altos insumos da matéria orgânica alóctone (produzida fora do lago) estimulam comunidades heterotróficas, como bactérias, zooplâncton e peixe, aumentando as taxas de respiração inteira. Portanto, é mais provável que pequenos lagos florestais sejam heterotróficos líquidos, com as taxas de ER excedendo as taxas primárias de produção no lago. Por outro lado, os lagos arborizados com baixa taxa de drenagem recebem relativamente menos nutrientes e matéria orgânica, geralmente resultando em lagos de água clara, com baixas taxas de GPP e ER (Tabela).

Outra diferença importante entre os lagos que influencia a variabilidade do metabolismo do lago é o tempo de permanência da água no sistema, especialmente entre os lagos que são intensamente gerenciados pelos seres humanos. Alterações no nível do lago e nas taxas de descarga afetam as concentrações de matéria orgânica e nutrientes, abundância de organismos e taxas de processos ecológicos, como a fotodegradação da matéria orgânica colorida, afetando as taxas metabólicas magnitudes e a variabilidade. Lagos ou lagos endoréicos com tempo intermediário de residência hidráulica (TRH) normalmente têm um alto tempo de retenção de nutrientes e matéria orgânica no sistema, que favorece o crescimento de produtores primários e a degradação bacteriana da matéria orgânica. Assim, espera -se que esses tipos de lagos mantenham taxas de GPP e ER relativamente mais altas e menos variáveis do que os lagos com baixo tempo de permanência no mesmo status trófico. Por outro lado, espera -se que os lagos com HRT longa tenham taxas metabólicas reduzidas devido a insumos mais baixos de nutrientes e matéria orgânica para o lago. Finalmente, os sistemas lênticos que têm mudanças frequentes e intensas no nível da água e as taxas de descarga acelerada têm uma dinâmica mais próxima dos sistemas lúdicos, com taxas geralmente baixas de GPP e ER, devido a nutrientes, matéria orgânica e algas sendo liberadas do sistema durante intensa Eventos de descarga.
Variação temporal em escala diária
Em uma escala diária, as taxas de GPP são mais afetadas pelo ciclo de diel da radiação fotossinteticamente ativa, enquanto o ER é amplamente afetado pelas mudanças na temperatura da água. Além disso, as taxas de ER também estão ligadas à quantidade ou qualidade do substrato orgânico e às contribuições relativas da respiração autotrófica e heterotrófica, conforme indicado por estudos dos padrões de respiração noturna (por exemplo, Sadro et al 2014). Por exemplo, a respiração do bacterioplâncton pode ser maior durante o dia e nas primeiras horas da noite, devido à maior disponibilidade de matéria orgânica dissolvida lábil produzida pelo fitoplâncton. À medida que o sol nasce, há um rápido aumento na produção primária no lago, geralmente tornando -o autotrófico (NEP> 0) e reduzindo o CO2 dissolvido que foi produzido a partir da mineralização de carbono que ocorreu durante a noite. Esse comportamento continua até atingir um pico na NEP, normalmente em torno da disponibilidade máxima de luz. Depois, há uma tendência para a NEP cair constantemente entre as horas de disponibilidade máxima de luz até o nascer do sol do dia seguinte.
As diferenças diárias na luz e na temperatura recebidas, devido a diferenças no clima, como cobertura de nuvens e tempestades, afetam as taxas de produção primária e, em menor grau, a respiração. Essas variações climáticas também causam variabilidade de curto prazo na profundidade da camada mista, que por sua vez afeta os nutrientes, matéria orgânica e disponibilidade de luz, bem como trocas de gás verticais e horizontais. A mistura profunda reduz a disponibilidade de luz, mas também aumenta os nutrientes e a disponibilidade de matéria orgânica nas camadas superiores. Assim, os efeitos da variabilidade de curto prazo na profundidade da camada mista na produção primária bruta (GPP) dependerão de quais são os fatores limitantes em cada lago em um determinado período. Assim, uma camada de mistura mais profunda pode aumentar ou diminuir as taxas de GPP, dependendo do equilíbrio entre a limitação de nutrientes e a luz da fotossíntese (Figura).
As respostas nas taxas metabólicas são tão dinâmicas quanto os processos físicos e químicos que ocorrem no lago, mas as alterações na biomassa de algas são menos variáveis, envolvendo crescimento e perda por períodos mais longos. A alta disponibilidade de luz e nutrientes está associada à formação de flores de algas em lagos; Durante essas flores, as taxas de GPP são muito altas e as taxas de ER geralmente aumentam quase tanto quanto as taxas de GPP, e o saldo de GPP e ER é próximo de 1. Logo após a flor, as taxas de GPP começam a diminuir, mas as taxas de ER continuam mais altas devido a A alta disponibilidade de matéria orgânica lábil, que pode levar a um rápido esgotamento da concentração de oxigênio dissolvida na coluna de água, resultando em mortes de peixes.
Variação temporal em uma escala anual
As variações sazonais no metabolismo podem ser impulsionadas por variações sazonais de temperatura, capa de gelo, precipitação, dinâmica de mistura e estratificação e sucessão comunitária (por exemplo, controle de fitoplâncton pelo zooplâncton). As variações sazonais no metabolismo do lago dependerão de como as estações alteram os insumos de nutrientes e matéria orgânica, e a disponibilidade de luz e dos quais os fatores estão limitando as taxas metabólicas em cada lago.
A luz é o principal fator do metabolismo do lago, portanto, a sazonalidade nos níveis de luz é um importante fator de mudanças sazonais nas taxas metabólicas do lago. Portanto, espera -se que as taxas de GPP sejam mais pronunciadas durante as estações como a primavera e o verão, nas quais os níveis de luz são mais altos e os dias são mais longos. Isso é especialmente pronunciado para lagos com GPP com limitação de luz, por exemplo, lagos mais turbidos ou manchados. A sazonalidade nos níveis de luz também afeta as taxas de ER. As taxas de respiração do ecossistema geralmente são acopladas às taxas de GPP, portanto, as estações com GPP mais altas também mostram taxas de ER mais altas associadas ao aumento da matéria orgânica produzida dentro do lago. Além disso, durante as estações com mais níveis de luz, a fotodegradação da matéria orgânica é mais pronunciada, o que estimula a degradação microbiana, aumentando as taxas de respiração heterotrófica.

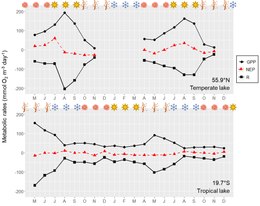
A maioria dos lagos do mundo congelou durante o inverno, um período de baixa irradiância, no qual a penetração da luz limite de cobertura de gelo e neve na coluna de água. A limitação da luz ocorre principalmente por cobertura de neve e não pelo gelo, o que torna a produção primária fortemente sensível à cobertura de neve nesses lagos. Além da limitação da luz, baixas temperaturas sob gelo também diminuem as taxas metabólicas, mas não o suficiente para interromper os processos metabólicos. Portanto, o equilíbrio metabólico é geralmente negativo durante a maior parte da estação do gelo, levando à depleção de oxigênio dissolvida. Lagos rasos em climas áridos não têm ou muito pouca cobertura de neve durante o inverno, portanto, a produção primária sustentada sub-gelo pode ser suficiente para evitar a depleção de oxigênio dissolvida, conforme relatado por Song e outros em um lago mongol. Apesar da alta proporção dos lagos mundiais que congelam durante o inverno, poucos estudos foram realizados no sub-gelo do metabolismo do lago, principalmente devido à amostragem de dificuldades técnicas. Os lagos que estão mais próximos do equador experimentam menos sazonalidade em relação à intensidade da luz e às horas da luz do dia do que os lagos em latitudes mais altas (zonas temperadas e polares). Assim, é mais provável que os lagos em latitudes mais altas experimentem limitação de luz da produção primária durante as estações de pouca luz (inverno e outono). As diferenças sazonais de temperatura também não são tão importantes nos trópicos como são para lagos de latitudes mais altos. Assim, o efeito direto das variações sazonais de temperatura nas taxas metabólicas é mais importante nos lagos de latitudes mais altos do que nos lagos tropicais (figura). Por sua vez, é mais provável que os lagos tropicais e subtropicais tenham variações sazonais após a dinâmica da estratificação e mistura e regimes de chuva (estações úmidas e secas), do que devido às quatro estações astronômicas ou meteorológicas (primavera, verão, outono e inverno) Quando comparado aos lagos mais altos de latitudes.

Mudanças sazonais de temperatura e chuva levam a mudanças sazonais na estabilidade da coluna de água. Durante períodos de estabilidade da coluna de baixa água, uma camada mista mais profunda (mistura total ou parcial da coluna de água, dependendo do lago) aumenta as entradas de nutrientes e matéria orgânica de camadas mais profundas e através da ressuspensão dos sedimentos, o que reduz a disponibilidade de luz. Por outro lado, durante períodos de estabilidade forte da coluna de água, são suprimidas cargas internas de nutrientes, matéria orgânica e bactérias associadas à coluna de água, enquanto a perda de algas devido ao afundamento é aprimorada. Além disso, a disponibilidade de luz durante esse período é maior, devido à fotodegradação, menor ressuspensão dos sedimentos e menor profundidade de mistura, que expõem o fitoplâncton a um ambiente mais rico em luz. Taxas de ER mais altas durante o período de estabilidade da coluna de baixa coluna de água, como conseqüência da maior disponibilidade de matéria orgânica e da maior biomassa de bactérias associadas a essa matéria orgânica, foram relatadas para muitos lagos em todo o mundo. No entanto, as respostas das taxas de produção primárias a essas mudanças sazonais demonstraram comportamentos diferentes em diferentes lagos. Como dito antes, as respostas das taxas metabólicas a essas mudanças dependerão de fatores limitantes da produção primária em cada lago (figura). Durante os períodos de estabilidade da coluna de água baixa, a ressurgência das águas ricas em nutrientes pode resultar em maiores taxas de GPP pelágicas, como foi observado em alguns lagos tropicais. Por outro lado, durante os períodos de estabilidade da coluna de água baixa, as taxas de GPP podem ser limitadas pela baixa disponibilidade da luz, como foram observadas em alguns lagos temperados e subtropicais. O equilíbrio metabólico líquido é geralmente mais negativo durante períodos descrorizados, mesmo em lagos em que a temporada bem misturada é o período mais produtivo. Independentemente da alta GPP nesses sistemas, as taxas de ER também são aprimoradas pelo aumento da disponibilidade de estoques de matéria orgânica de sedimentos e águas mais profundas.
As diferenças sazonais nas chuvas também afetam as taxas metabólicas. O aumento da precipitação promove a entrada de matéria e nutrientes orgânicos nos lagos, o que pode estimular as taxas de ER e estimular ou inibir as taxas de GPP, dependendo do equilíbrio entre o aumento dos nutrientes e a menor disponibilidade da luz. Por outro lado, a menor precipitação também afeta as condições limnológicas, reduzindo o nível da água e, assim, aumentando a concentração de nutrientes e a clorofila, além de alterar a estabilidade térmica dos ambientes aquáticos. Essas mudanças também podem melhorar as taxas de ER e GPP. Assim, o grau das respostas das taxas metabólicas às mudanças sazonais nas chuvas dependerá da morfometria do lago, propriedades de captação e a intensidade e duração dos eventos de chuva. Os lagos freqüentemente expostos a tempestades fortes, como as áreas do tufão no noroeste do Oceano Pacífico, recebem eventos intensos de chuva que podem durar alguns dias. Durante essas temporadas de tempestade, é esperada uma redução nas taxas metabólicas devido à redução da luz solar e à descarga de água e organismos. Espera -se que essa redução seja mais pronunciada no GPP do que nas taxas de ER, resultando em um NEP mais heterotrófico (GPP
Variações interanuais
A variabilidade interanual nas taxas metabólicas pode ser impulsionada por extensas mudanças na bacia hidrográfica ou por mudanças climáticas direcionais e cíclicas e distúrbios climáticos, como os eventos associados à oscilação do sul do El Niño (ENSO). Essas mudanças na bacia hidrográfica, a temperatura do ar e a precipitação entre os anos afetam as taxas metabólicas, alterando insumos de nutrientes e matéria orgânica para o lago, atenuação da luz, dinâmica de mistura e por dependência direta de temperatura de processos metabólicos.
O aumento da precipitação aumenta a carga externa de matéria orgânica, nutrientes e sedimentos nos lagos. Além disso, o aumento dos eventos de alta promovido pelo aumento das chuvas também pode alterar a dinâmica de mistura e causar descarga física de organismos. Enquanto a menor precipitação associada a altas taxas de evaporação também afeta as condições limnológicas, reduzindo o nível da água e aumentando assim a concentração de nutrientes e a clorofila, além de alterar a estabilidade térmica dos ambientes aquáticos. Durante os anos mais quentes, uma estabilidade mais forte da coluna de água limita as entradas de nutrientes e matéria orgânica para a zona fótica. Por outro lado, durante anos mais frios, uma coluna de água menos estável aumenta a ressuspensão dos sedimentos e as entradas de nutrientes e matéria orgânica de águas mais profundas. Isso diminui a disponibilidade de luz, enquanto aumenta a disponibilidade de nutrientes e matéria orgânica. Assim, os efeitos das diferenças na precipitação e temperatura entre os anos nas taxas metabólicas dependerão da intensidade e duração dessas alterações, e também nos quais os fatores estão limitando o GPP e o ER em cada corpo de água.
Em lagos com nutrientes e limitação de matéria orgânica de GPP e ER, os anos mais úmidos podem aumentar as taxas de GPP e ER, devido a insumos mais altos de nutrientes e matéria orgânica da paisagem. Isso dependerá se os insumos terrestres estarão prontamente disponíveis para os principais produtores e comunidades heterotróficas ou se entrará no lago através de águas mais profundas, nas quais os processos metabólicos são muito baixos ou inexistentes. Nesse caso, as entradas estarão disponíveis apenas no próximo evento de mistura de coluna de água. Assim, aumentos nas taxas metabólicas devido à precipitação dependem também da estratificação e dinâmica de mistura, hidrologia e morfometria do lago. Por outro lado, os anos mais secos também podem ter taxas de GPP e ER aprimoradas se forem acompanhadas por níveis mais baixos de água, o que levaria a nutrientes mais altos e concentrações de matéria orgânica. Um nível de água mais baixo está associado a uma coluna de água menos estável e maior proximidade com os sedimentos, aumentou assim os insumos de nutrientes e matéria orgânica de águas mais profundas. Além disso, uma redução no nível da água através da evaporação da água leva a um efeito de concentração. Por sua vez, durante os anos mais quentes, a coluna de água é mais estável e a profundidade da camada de mistura é mais rasa, reduzindo assim as entradas internas de nutrientes e matéria orgânica para a camada mista. As taxas metabólicas, nesse cenário, serão mais baixas na camada mista superior. Nos lagos com uma zona fóica que se estende mais profunda que a camada mista, as taxas metabólicas serão mais altas em profundidades intermediárias, coincidindo com os máximos de clorofila profunda.
Nos lagos com produção primária limitada principalmente pela disponibilidade de luz, os aumentos nas chuvas podem levar a menor disponibilidade da luz, associada ao aumento da matéria orgânica dissolvida e à matéria total suspensa. Consequentemente, o aumento das chuvas estaria associado a níveis mais baixos de GPP, o que reduziria as taxas de respiração associadas à produção autóctone, levando a uma dissociação das taxas de GPP e ER. Além disso, o aumento da disponibilidade de matéria orgânica alóctones durante os anos úmidos pode levar a ER mais alta e, consequentemente, levar o equilíbrio metabólico a ser negativo (NEP <0).
Alterações na precipitação anual também podem afetar a variabilidade espacial nas taxas metabólicas nos lagos. Williamson e colaboradores, por exemplo, descobriram que, em um reservatório hiper-eutrófico na América do Norte, a variabilidade espacial relativa nas taxas de GPP e ER foi maior em um ano seco em comparação com um molhado. Isso sugere maior relevância de processos internos, como carregamento interno, captação de nutrientes, sedimentação e ressuspensão, a taxas metabólicas durante anos secos.